
Drylands are fragile ecosystems constantly enduring polyextreme conditions (e.g. few nutrients, high UV irradiation, significant daily and seasonal temperature fluctuations) and, by definition, present an overall shortage of bioavailable water.
These lands cover more than 45% of the Earth’s surface, are home to ~3 billion humans (~37.5% of the population) and generate ~50% of global food production. However, ongoing climate change (including rising temperatures) entails aridity intensification, leading to the worldwide expansion of dryland surfaces and the disruption of the existing ones, a process known as desertification.
The protection and restoration of drylands are ecologically, socially, and economically crucial. With ongoing climate changes, drylands are expanding and becoming drier. Desertification threatens 250 million human beings and 70% of actual drylands, and it will put 1 billion more human beings at risk of famine within the next decade, with nearly 50 million people likely to be displaced because of the loss of biodiversity, soil fertility and productivity. For this reason, we need to modify our perspective on drylands and act now, revising the strategies for their protection with the use and application of ‘microbe-based solutions’.
Microbial communities are fundamental (and well-adapted) components of dryland ecosystems, as they provide most of the essential ecosystem services that support the resilience of these soils/lands. At the same time, through their long-evolved intimate relationships with plants, microorganisms can support and promote vegetation survival and adaptation. However, microorganisms are usually ignored in global-scale desertification mitigation strategies. Following Albert Einstein’s maxim “a problem without a solution is a poorly stated problem”, we advocated for the use of dryland microorganisms to (i) alleviate soil erosion and enhance soil fertility/stability (e.g. preservation and promotion of biological soil crusts), (ii) promote revegetation, and (iii) even improve desert farming production. All these strategies can be part of a framework where microbial-based solutions are included at the forefront of aridification control actions (i.e. reduce, avoid, and reverse) in drylands.

Microbial ecology and function within dryland environments
Climate changes, and in particular global warming and extended drought periods, are causing shifts in the distribution, diversity, abundance, and functionality of microbial species, influencing microorganism–microorganism and plant/soil–microorganism interactions with uncertain and unknown consequences on the overall metabolic capabilities of terrestrial ecosystems.
Like any living cell, microorganisms are essentially composed of water, which allows intracellular diffusion of (macro)molecules, such as proteins and metabolites, but is also crucial for many reactions, such as photosynthesis. However, as the name suggests, drylands are ecosystems suffering from global water shortage, which already imposes strong selective pressure on their indigenous communities. The further aridification of (hot) dryland ecosystems caused by global climate change will lead to dramatic, and maybe irreversible, changes in their functioning. Microbial communities are key to dryland global functioning as they are important ecosystem service providers (e.g., primary production). Furthermore, some of these microorganisms may have already reached the upper limit of aridity tolerance. With water availability further reduced, we can expect decreasing microbial diversity, biomass and capacity to provide ecosystem services, as only a fraction of the original communities will be able to cope with the new water stress condition. The research performed by Jean-Baptiste Ramond’s group aims at better understanding the adaptation of environmental microbial communities to extreme environmental conditions, particularly in the hyperarid Namib and Atacama deserts, as well as to climate change. This entails the study of the diversity and functional capacities of microbial communities from desert soils and refuge niches/microbial hotspots, such as lithic environments, using in situ and climate change-mimicking controlled experiments.
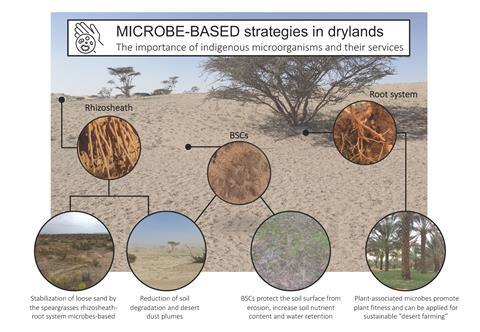
This approach is also used by Federico Rossi and Marc W. Van Goethem to evaluate the role of edaphic microorganisms in the formation/stabilisation of Biological Soil Crust (BSCs), essential photosynthetic microbial communities at the soil surface in many arid and semiarid environments. Special scientific attention has been devoted to cyanobacteria, a dominant and essential fraction of the BSC community. These special prokaryotes release chemically complex extracellular polymeric substances and various biomolecules that profoundly affect biological activity and soil productivity. Indeed, along with the study of BSC composition, F. Rossi explores (and exploits) the possibility of using cyanobacteria as soil bioinoculants as part of land restoration strategies. An effective inoculation of cyanobacteria can lead to the formation of induced biological crusts, with proven benefits in terms of water availability at the soil surface and soil stability against erosion.

At both laboratory and field scales, F. Rossi, in collaboration with American and Chinese research groups, studies the contribution of BSCs and their associated extracellular polymeric substances in soil hydrological cycles and soil multifunctionality, to identify factors (environmental and biogenic) that positively or negatively affect the establishment of cyanobacteria at the inoculation site to optimise the inoculation protocols. For instance, by studying the effects of changing climate on BSCs, M. W. Van Goethem showed that during the rewetting of BSCs in lab conditions, the expected bloom of Firmicutes (Bacillus) triggers the activation of specific phages (Caudovirales) that control their abundance throughout the year, consistently with the classical ‘kill-the-winner’ hypothesis.
Along with BSCs, vegetation and their associated root systems represent a unique ‘micro-oasis’ in which edaphic microorganisms proliferate. Indeed, the rhizosphere, the narrow zone of soil around plant roots, is characterised by a high concentration of microorganisms, including bacteria, fungi and archaea. As studied by Ramona Marasco and Daniele Daffonchio in different extreme/arid ecosystems, plant-associated microorganisms play important roles in plant growth and ecosystem function and soil fertility and edaphic-niche stability. Such biodiversity/soil multifunctionality mediated by microorganisms plays a crucial role in restoring degraded soils and combating desertification, underlining the importance of nature-based solutions to counteract the negative effects of climate changes on drylands. However, it is important to investigate the possible effective strategies to apply nature-based solutions, and.,ll ‘omics’ could give important insights. As reviewed by M. W. Van Goethem, during the last century, research on microbial communities in drylands took advantage of omics to expand our knowledge of their composition, ecology and functional role. These novel methods and approaches could also help in disentangling the abiotic and biotic factors shaping the microbial community in the ecosystem, revealing a plethora of biologically mediated environmental processes, such as secondary-metabolite biosynthetic potential and biosynthetic gene clusters (BGCs) encoding secondary metabolites.
On the scale of ecological catastrophes, dryland conservation and restoration are more urgent than monitoring off-target effects. In this context, our opinion promotes the concept/idea that we can no longer “wait and see” but that it is now the time to act and make decisions. We also want to underline that along with the need to act, the ongoing (and planned) mitigation/restoration strategies cannot afford to neglect the entire dryland ecosystem, including dryland soils and their associated microorganisms.
The knowledge and data generated by all of the researchers above-mentioned ultimately aim at improving climate change modelling with microbial data and designing microbial-based strategies that could be used to improve plant (including food crop) growth and productivity in more arid conditions and fight desertification.
We argue that dryland microbial communities should be the cornerstone of ‘microbe-based solutions’ for any environmental initiative strategy designed to mitigate the effects of climate change in drylands. Such a bio-based strategy will unequivocally help to conserve/sustain/promote dryland diversity and food production and give microorganisms-as we are microbiologists-the positive attention they deserve.
Ramona’s latest research on dryland ecosystems is out in Microbial Biotechnology and can be accessed here.






No comments yet