
Do you work in forensic microbiology, or have an interest in it? Click here to find out about our upcoming AMI event on the topic! This article was originally written and published in 2015. Therefore, certain wording may reflect this.
Almost in the fashion of the legendary phoenix, microbial life springs from decaying matter such as the carcass of an animal. The decomposition process is driven by bacteria, and these microbes not only spread across the carrion, but gather together forming biofilms. In both cases, cell signalling between bacteria plays an important role as does, inter-kingdom signalling between bacteria and insects. By understanding the temporal events related to the decomposition of an animal resource, the time of death may be approximated.
Cell signalling
Many bacteria communicate by constantly secreting small molecules, as the number of bacteria increases, the concentration of these compounds increases until a threshold concentration is reached. The signal is then internalised or activates a membrane receptor that leads to changes in gene expression and increased production of the original signal. This method of communication is known as quorum sensing (QS) and using QS, bacteria sense their numbers and begin to act in concert. This leads to a change from single cell behaviour to group behaviour and the cells resemble primitive tissues. QS is particularly important for the expression of virulence traits for many pathogens, and it is important for the rapid movement across surfaces known as swarming. Other traits such as competence, conjugation, antibiotic production, sporulation, and biofilm formation are also influenced by QS. Interestingly, often the ability of the bacterium to respond to environmental stress is also linked to the QS ability of the cells. In addition to bacterial cell signalling, bacteria also communicate with their surroundings across kingdoms using chemicals cues, influencing animals such as insects during their utilisation of an animal resource.
Swarming
One manifestation of group behaviour based on cell signalling is the spreading of bacteria over the animal resource via swarming; the cells move to take complete advantage of the resources presented by the decaying animal. In swarming, cells become hyper-flagellated and walk on the surface of the carrion in pulsed rounds of rapid movement that depend on bacterial signalling (Fig. 1). To facilitate swarming, cells frequently secrete glycolipid or lipopeptide biosurfactants as wetting agents to reduce surface tension. In this way, bacteria like Proteus mirabilis rapidly cover the surface of the decaying animal.
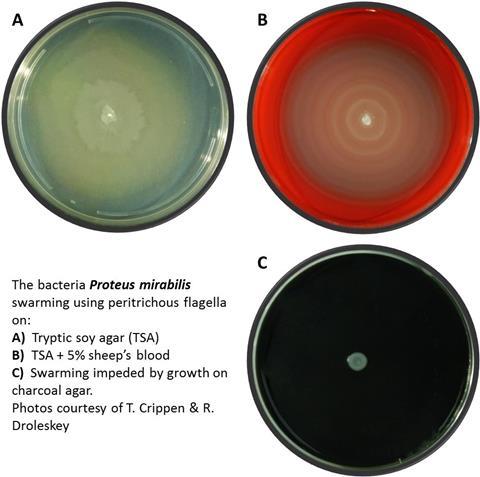
Biofilm formation and dispersal
Not all the cells choose to travel. As the bacteria spread, they also form biofilms, vibrant communities of cells (Fig. 2) where cells are cemented together by protein and sugar polymers and even by DNA (a fine example of nucleotides serving a structural rather than a genetic role). By living in biofilms, cells are protected from changes in the environment, and they can devour the decaying animal.
To commence biofilm formation, bacteria attach to surfaces via their appendages, such as fimbriae and flagella, and microcolonies begin to form as cells cement themselves together in what has been termed the biofilm matrix. For some strains, pioneering cells leave trails of matrix sugar polymers and DNA which attract other cells to the growing biofilm and create connecting channels. Eventually, cells may build architectural marvels that allow for nutrients to enter and waste to be removed.
The key internal switch that converts motile bacteria to sessile ones (and back to motile ones) is cyclic diguanylate (c-di-GMP): in all bacteria examined to date, biofilm formation increases as c-di-GMP levels increase. In an elegant regulatory balance, often involving many forms of each of these types of enzymes, cells use diguanylate cyclases to form c-di-GMP and phosphodiesterases to reduce c-di-GMP levels. Overall, c-di-GMP levels are first low to increase motility to the surface, high to form biofilms and reduce motility, then low again to allow cells to become motile again. In effect, a form of guanosine, the building block of RNA, is converted into an internal signal that is used by the cells to control biofilm formation (another example of cells using nucleotides in a role not directly related to genes).
As with us, cells in biofilms often leave their homes, this behaviour is termed biofilm dispersal. Dispersal allows cells to leave in search of more nutrients as well as to colonise more locations when nutrients are plentiful. For example, the opportunistic pathogen Pseudomonas aeruginosa will disperse in response to both a sudden decrease and a sudden increase in nutrients. Dispersal requires cells to remove the cement that tethers them together and often to become motile. Internally, the signal for biofilm dispersal is a reduction in c-di-GMP levels brought on by the activation of phosphodiesterases.

Inter-kingdom signalling on carcasses
Cell signalling, swarming, and biofilm formation/dispersal are all utilised to allow bacteria to recycle carrion. Some of the volatile signals of decay that are produced by the bacteria are perceived by animals such as blow flies. These signals are derived from the non-odorous proteins, fats, and oils of the animal resource. Ammonia is the most common nitrogenous product and is formed as a secondary product of urea decomposition. Skatole, indole, mercaptans, and sulphides make the most penetrating odours of putrefaction. Another large group of attractants are fatty acids, which are usually bacterial fermentation products and decomposition components.
A connection has been found between the signals used by bacteria for swarming over the animal resource and for attracting flies that help transport the bacteria to new resources; hence, signalling between bacterial cells and between bacteria and flies are interrelated. By using a bacterium, Proteus mirabilis, derived from the salivary glands of a blow fly, Lucilla sericata transposon knockouts were created and the mutants were screened for reduced swarming. It was found that fly attractants such as putrescine, lactic acid, phenol, NaOH, KOH, and ammonia also restore swarming for cells with the swarming mutations. In addition, cells deficient in swarming attracted fewer blow flies and reduced their egg laying. Hence, well-known fly attractants also facilitate bacterial swarming. Therefore, as the bacteria swarm they attract flies, with their volatile swarming signals, which then transport them to new resources, and they alert flies to lay eggs to help the decay process.
Indole plays a prominent role in inter-kingdom signalling between bacteria and insects that facilitate carrion decay. Indole is a QS compound for E. coli (this bacterium secretes mM quantities of this compound), and in the gastrointestinal tract, indole from E. coli both tightens the host epithelial cell junctions as well as reduces the virulence of pathogens such as enterohaemorrhagic E. coli and P. aeruginosa. For insects, indole has been isolated from human skin as a bacterial metabolite that elicits a strong response by mosquitoes, it also attracts blow flies.
Biofilm forensics
Since bacteria initiate the decay of the animal resource and their volatile degradation products attract insects, the timing of death should be related to the extent of bacterial swarming, the extent of biofilm formation, and the extent and type of fly egg laying. Since different flies have different olfactory preferences, it should be possible to relate the timing of the death to the volatiles produced as well as to the type of insect that is attracted. Therefore, understanding the progression of carrion decay in terms of the degree of bacterial swarming, the volatile signals produced by the bacteria, and the extent and type of insect colonisation should lead to more accurate predictions of the demise of the animal.






No comments yet